The Invention of Sexual Reproduction
The evolutionary game-changer that supercharged genetic diversity.
The Shuffle That Made Death Worth It
Sex is older and stranger than romance lets us believe. Long before bodies, courtship, or even animals, single cells learned to do something no chemistry had ever required: fuse two genomes, halve them through meiosis, and pass on a child that resembled neither parent exactly. The Invention of Sexual Reproduction was not the invention of pleasure or even of two genders. It was the invention of recombination as a way of life — and it quietly rewrote the rules for everything that came after, including us.
The Preconditions: A Cell Complex Enough to Risk It
Sex needed a stage. That stage was the eukaryotic cell, with its nucleus, its packaged chromosomes, and its capacity for the elaborate chromosomal choreography of meiosis. So this event sits directly downstream of The First Complex Cells (Eukaryotes) (sv-first-complex-cells), itself a product of the oxygen-rich world opened by The Great Oxygenation Event (sv-great-oxygenation), which in turn traces back to the deeper miracle of The Origin of Life (sv-origin-of-life). Reconstructions of the last eukaryotic common ancestor (LECA) suggest it was already sexual — meaning the fusion of haploid gametes and meiotic recombination were near-foundational to the eukaryotic lineage, not a later luxury. The same atoms that made all of this — the carbon and oxygen forged in stars and scattered by The First Supernovas (sv-first-supernova) — were now being shuffled in a wholly new way.
Why Pay the Cost?
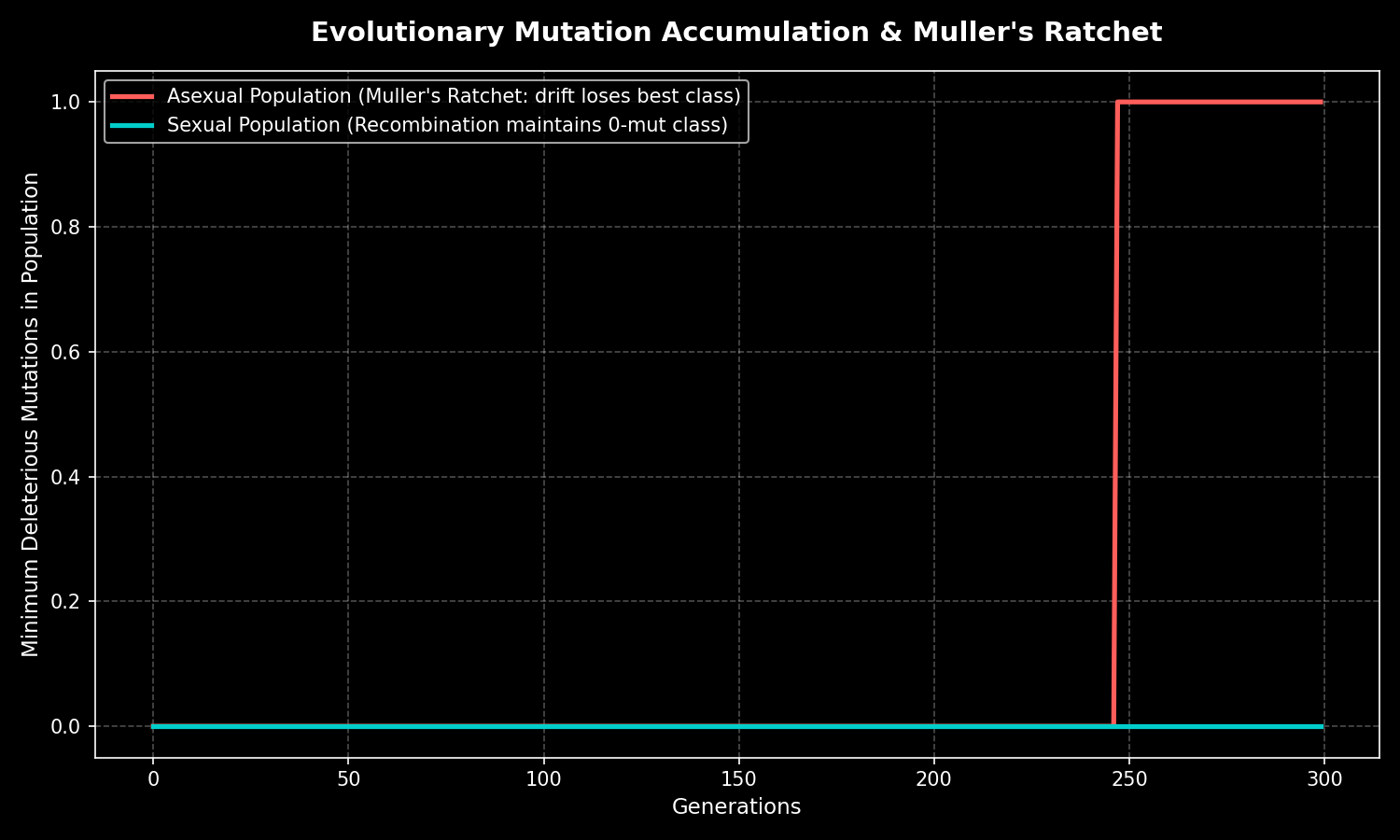
Sex is expensive. An asexual lineage clones itself wholesale; a sexual one throws away half its genome each generation and must find a partner. Biologists have spent a century asking why evolution tolerated that "twofold cost." The leading answers are sobering and beautiful. Muller's ratchet holds that asexual genomes silently accumulate deleterious mutations they can never undo; recombination lets sexual lineages purge that load. The Red Queen hypothesis frames sex as an arms race — by reshuffling genes each generation, hosts produce rare genotypes that coevolving parasites cannot pre-adapt to. Sex, in this reading, is not about making more offspring but about making unpredictable ones. Variation became a defense, and a fuel.
The Ripple: Variation as an Engine
That fuel transformed history's tempo. By generating combinatorial diversity each generation, sexual reproduction handed natural selection an endlessly reshuffled deck. The Cambrian Explosion (sv-cambrian-explosion) — the sudden flowering of animal body plans, foreshadowed by The Ediacaran Biota (sv-ediacaran-biota) — is hard to imagine without the variational engine that sex provides. Every branching event since runs through it: the diversification of The Evolution of Sharks (sv-first-sharks), the conquest of land at Tiktaalik & The Move to Land (sv-tiktaalik), the patient rise of The First True Mammals (sv-first-mammals). When Darwin built his theory in The Origin of Species (sv-charles-darwin), the raw material he assumed — heritable variation — was largely sex's gift, though he never knew the molecular machinery beneath it.
The Bargain With Death
Sex also bound life to mortality in a new way. Cloning is, in principle, a kind of immortality; the sexual organism, by contrast, is a temporary recombination meant to be superseded. We are, biochemically, vehicles built to mix genomes once and step aside. In that sense this ancient cellular event still shapes the most modern human anxieties — the longevity dreams of Longevity Escape Velocity (sv-kurzweil-lev) and the vision in Biology Becomes Information Technology (sv-kurzweil-genome) are, at bottom, attempts to renegotiate a bargain a microbe struck billions of years ago. The shuffle that made us creative also made us finite. Nearly every story this timeline tells is, distantly, the story of two cells deciding to merge.
Global Context
Sexual reproduction is not datable to a calendar moment but to a deep window in the Proterozoic Eon. The genetic machinery of meiosis (SPO11, DMC1, the MutS/MutL homologs) is conserved across all eukaryotic lineages, implying the last eukaryotic common ancestor (LECA), perhaps 1.8–2.0 billion years ago, was already sexual. This places sex in the wake of two prior revolutions: the Great Oxidation Event (~2.4 Ga), which oxygenated the atmosphere, and the endosymbiotic acquisition of the mitochondrion, which made the energetic, internally complex eukaryotic cell possible. The oldest direct fossil evidence comes much later: Nicholas Butterfield's Bangiomorpha pubescens from Somerset Island, Nunavut, a bangiophyte red alga preserving differentiated spore-bearing reproductive structures, redated by rhenium-osmium methods to roughly 1.047 billion years. Beyond this microbial-to-algal world there were no animals, no plants, no land life. Earth was a "boring billion" of stable oceans dominated by prokaryotes and single-celled eukaryotes, with sex quietly establishing the genetic engine that the Cambrian explosion would later exploit.
The Paradigm Shift
Sex fused two innovations: meiosis, which halves the genome and reshuffles it via recombination, and syngamy, the fusion of gametes to restore ploidy. The consequence was a permanent mechanism for generating genetic variation within populations rather than relying solely on mutation. Recombination unlinks alleles, letting natural selection act on them independently; it builds favorable gene combinations and purges deleterious ones that asexual lineages accumulate irreversibly (Muller's ratchet). This transformed the tempo and mode of evolution: sexual populations could track changing environments and coevolving parasites far faster than clones. The downstream redirection is profound. Anisogamy (the divergence of large eggs and small sperm) created two sexes, the substrate for sexual selection that Darwin invoked to explain peacocks and antlers. Recombination underwrote the diversification of multicellular eukaryotes, ultimately the Cambrian radiation. Genealogically, sex made "species" coherent reproductive communities and gave biology its tree-and-network structure. Nearly every macroscopic organism alive descends from this Proterozoic invention; it is the genetic precondition for complex life as we know it.
Counterfactual: What If It Had Gone Differently
Had eukaryotes remained obligately asexual, evolutionary theory predicts a markedly impoverished biosphere. Without recombination, lineages accumulate deleterious mutations irreversibly through Muller's ratchet and suffer clonal interference, in which beneficial mutations in separate individuals cannot be combined and instead compete. Asexual clades typically appear as evolutionary "twigs"—young and short-lived—because they fail to purge mutational load or adapt to fast-coevolving parasites (the Red Queen dynamic of Lively and Hamilton). The rare persistent exceptions, the bdelloid rotifers that Maynard Smith dubbed an "evolutionary scandal," apparently survive via desiccation-driven horizontal gene transfer and DNA repair, surrogates for the variation sex provides. A wholly asexual eukaryotic world would plausibly have stalled at the level of slowly diversifying microbes and simple multicellular forms, with little of the rapid adaptive radiation seen in the Phanerozoic. The Cambrian explosion, sexual selection, and the genetic interchange that defines species would be absent or radically attenuated. Whether complex animal life could arise at all without sex remains genuinely uncertain and debated.
Scholarly Debate
The central, still-unresolved debate is the "queen of evolutionary biology" problem: why sex is maintained despite the twofold cost of producing males that John Maynard Smith and George C. Williams sharpened in the 1970s. Competing, non-exclusive hypotheses persist. The Mutational Deterministic / Muller's ratchet school (Alexey Kondrashov; Maynard Smith) holds that recombination's chief value is purging deleterious mutations. The Red Queen camp (W. D. Hamilton, Curtis Lively, with experimental support from Lively's Potamopyrgus snails and Levi Morran's Caenorhabditis work) argues coevolving parasites favor genetic novelty. Others emphasize the Fisher-Muller advantage of combining beneficial mutations. A separate dispute concerns origins versus maintenance—why sex arose at all—where some (Lynn Margulis controversially) tied it to cannibalism and DNA repair, while others stress meiosis as derived from ancient recombinational repair machinery. Recent work (Lane, Radzvilavicius) links the origin of two sexes to selection for mitochondrial quality. The fossil dating of Bangiomorpha itself was revised, and the inference that LECA was sexual, though now consensus, rests on molecular phylogenetics rather than direct evidence.
How It Connects
What Made It Possible
- The endosymbiotic origin of the eukaryotic cell roughly 1.5-2 billion years ago, in which an archaeal host acquired an alpha-proteobacterial endosymbiont that became the mitochondrion, created the membrane-bound nucleus and bioenergetic complexity that meiotic sex required as a platform.
- Prokaryotes had already evolved homologous-recombination machinery for DNA repair, including the RecA recombinase in bacteria and RadA in archaea, whose direct descendants Rad51 and Dmc1 catalyze the strand-exchange step of eukaryotic meiosis.
- Bacterial natural transformation—the uptake of environmental DNA and its integration via RecA-mediated strand invasion—provided the evolutionary template from which meiotic recombination is thought to have been derived in the earliest single-celled eukaryotes.
- The constant pressure of DNA damage (double-strand breaks in particular) selected for double-strand-break repair using a homologous template, the process Harris Bernstein and colleagues argue was repurposed into meiotic recombination so that a homologous partner chromosome could template repair.
- The evolution of multiple linear eukaryotic chromosomes, organized with centromeres and telomeres, supplied the substrate for homologous pairing and orderly segregation—linear chromosomes are regarded as a prerequisite for productive meiosis.
- Genome expansion and the spread of repetitive sequences in early eukaryotes limited the effectiveness of one-gene-at-a-time lateral gene transfer, favoring a switch to whole-genome meiotic recombination as the means of shuffling genetic information between individuals.
Its Legacy
- Because sex arose once in the last eukaryotic common ancestor and is shared across nearly all eukaryotic lineages, it became the default reproductive mode for the overwhelming majority of plants, animals, fungi, and protists alive today.
- Genetic recombination let lineages combine beneficial mutations from different individuals (the Fisher-Muller effect) and purge deleterious ones, halting the irreversible accumulation of harmful mutations that Muller's ratchet inflicts on asexual populations.
- Recombination supplied the continually reshuffled genetic variation that powers the Red Queen dynamic, allowing host populations to keep generating novel genotypes fast enough to resist co-evolving parasites and pathogens.
- The evolution of anisogamy—the split into small, numerous male gametes and large, costly female gametes—created the two sexes and, under the Darwin-Bateman paradigm, set the asymmetry that drives sexual selection, mate competition, and sex-specific traits.
- By accelerating adaptation and generating heritable variation, sexual reproduction raised speciation rates and fueled the adaptive radiations that produced much of Earth's biodiversity, with simulation studies finding higher speciation in sexual than asexual lineages.
- Sex was an early eukaryotic innovation tightly associated with the repeated origins of complex multicellularity, helping shape the large-bodied plants, animals, and fungi across the Tree of Life.
Myth vs. Reality
Myth: Sex evolved so that species could create more genetic variation and survive better — it's 'good for the species.'
Reality: This is a discredited group-selection argument. Modern evolutionary biology holds that traits spread because they benefit the individuals (or genes) carrying them, not because they benefit a species as a whole. As reviewers of the group-selection debate note, the older claim that a trait 'evolved because it was good for the species' is regarded as misleading; species-level selection is rejected by nearly all biologists. The persistence of sex is instead explained by individual-level advantages such as the Red Queen dynamic (out-evolving parasites) and the purging of harmful mutations — and even these remain actively debated, because sex carries a real individual-level 'twofold cost.'
Myth: Sexual reproduction has existed roughly since life began, billions of years before complex life.
Reality: Sex is a eukaryotic innovation, not a property of the earliest life. The molecular machinery of meiosis is shared across all eukaryotic lineages, which leads scholars to conclude that sex arose just once, in the last eukaryotic common ancestor (LECA), on the order of 2 billion years ago. Life itself, however, is far older (around 3.5–4 billion years) and spent its first billion-plus years as bacteria and archaea reproducing asexually. So sex is a relatively late arrival, tied specifically to the eukaryotic cell.
Myth: The first sexual organisms had males and females.
Reality: The earliest sexual eukaryotes were single-celled and isogamous — they fused two gametes of equal size, so there were no 'males' or 'females' at all. Distinct sexes arise only with anisogamy (gametes of unequal size: small 'male' sperm vs. large 'female' eggs), which evolved later from isogamy. Anisogamy is thought to have evolved independently many times across different lineages (land plants, various algae, malaria parasites, animals). Males and females are therefore a downstream consequence of sex, not a precondition for it.
Myth: Bacteria reproduce sexually — bacterial conjugation is just the microbial version of sex.
Reality: Bacterial conjugation, transformation, and transduction transfer DNA between cells, but they are not sexual reproduction in the eukaryotic sense. Conjugation typically moves only a portion of one cell's genome one-way into a recipient and produces no new offspring — an existing cell is modified, not created. True sex involves meiosis, the alternation of haploid and diploid states, and the union of complete genomes from two parents via gametes — none of which occurs in bacteria. These bacterial processes are better described as horizontal gene transfer or 'parasex.'
Myth: Meiosis and sex must have appeared together as a single sudden package.
Reality: Several researchers argue the components assembled gradually rather than all at once. Work by Adam Wilkins and Robin Holliday proposed that key steps of meiosis (such as the tight pairing of homologous chromosomes enabling safe recombination) predate full-blown sexual reproduction. More broadly, meiotic sex is thought to have evolved out of the pre-existing recombination/lateral-gene-transfer machinery inherited from archaea and bacteria, with the expansion of early eukaryotic genomes and proliferation of repeated sequences favoring the shift from lateral gene transfer to organized meiotic recombination. The picture is one of stepwise tinkering with older parts, not a single leap.
In Their Words
"I have become increasingly bothered by a major inconsistency between one conspicuous aspect of the biota and what I understand to be the theory of natural selection... the prevalence of sexual reproduction in higher plants and animals is inconsistent with current evolutionary theory... there is a kind of crisis at hand in evolutionary biology." — George C. Williams, Sex and Evolution (Princeton University Press, 1975), Preface/Chapter 1
Data Visualization
References & Sources
- George C. Williams, Sex and Evolution (Princeton University Press, 1975)
- John Maynard Smith, The Evolution of Sex (Cambridge University Press, 1978)
- Nicholas J. Butterfield, 'Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes,' Paleobiology 26 (2000)
- Curtis M. Lively & W. D. Hamilton; and Levi T. Morran et al., 'Running with the Red Queen: Host-Parasite Coevolution Selects for Biparental Sex,' Science 333 (2011)
- Nick Lane, The Vital Question / Arunas L. Radzvilavicius & Nick Lane on mitochondrial selection and the origin of sexes