The First Complex Cells (Eukaryotes)
One cell swallowed another, and accidentally invented all complex life.
The Merger That Made Complexity Possible
For roughly two billion years after life began, Earth was a planet of prokaryotes — bacteria and archaea, cells of breathtaking metabolic ingenuity but stubborn structural simplicity. The origin of the eukaryote, the complex cell that builds every animal, plant, fungus, and protist, was not the slow accumulation of small improvements but a singular, improbable union. Somewhere between roughly 2.2 and 1.5 billion years ago, an archaeal host cell — most likely a member of the recently discovered Asgard lineage — engulfed or merged with a free-living bacterium related to modern Alphaproteobacteria. That captive became the mitochondrion, and with it the deepest threshold in the history of life was crossed.
Deep Preconditions
The eukaryotic merger was a child of everything that came before it. It required the heavy elements forged in the first stars (sv-first-stars) and scattered by the first supernovas (sv-first-supernova), the watery, mineral-rich world assembled at the formation of the Earth (sv-earth-formation), and the chemistry that produced the origin of life (sv-origin-of-life) itself. Most directly, it leaned on the atmospheric transformation of the Great Oxygenation Event (sv-great-oxygenation). The engulfed bacterium specialized in oxidative respiration, the high-yield metabolism that oxygen makes possible. Molecular-clock estimates increasingly cluster the eukaryotic origin in the window after oxygen's rise, a synchrony many researchers read as causal rather than coincidental.
Why It Mattered: The Energetics of Complexity
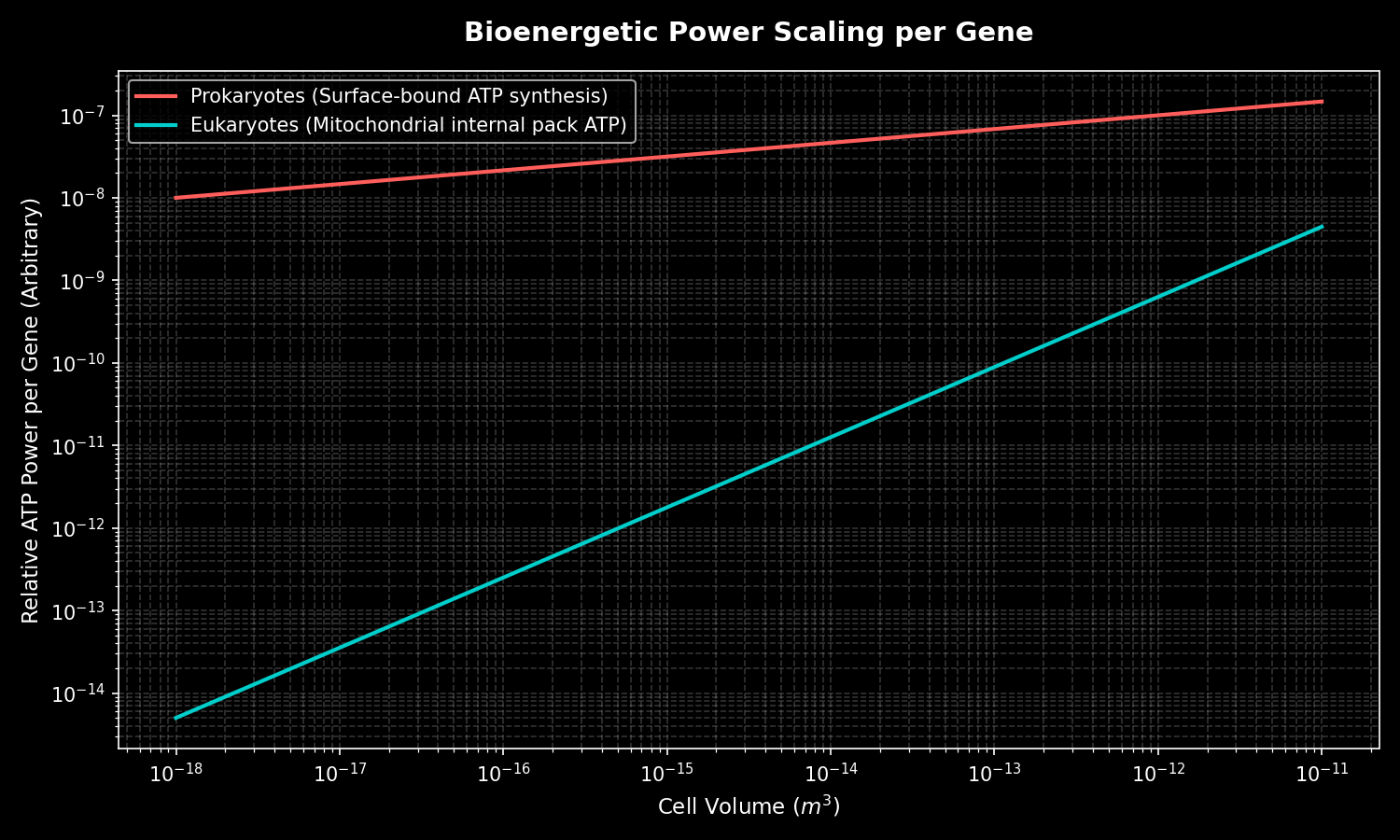
The biologist Nick Lane and the evolutionary biochemist William Martin argued in 2010 that mitochondria did something no prokaryote could do alone: they internalized energy production across vast areas of folded membrane, granting the cell orders of magnitude more available energy per gene. By their reckoning, a eukaryotic cell can support a genome thousands of times larger than a bacterium's. Prokaryotes, they argued, are trapped at the surface of their own cell membranes, energetically forbidden from expanding their genomes. Eukaryotes, powered from within by hundreds or thousands of mitochondria, escaped that ceiling. This is why complexity — large genomes, nuclei, internal compartments, the eventual leap to multicellularity — is overwhelmingly a eukaryotic story. The merger did not merely add a feature; it rewrote the rules of what a cell could afford to become.
The Ripple Forward
Almost everything visible in the later timeline descends from this one event. The eukaryotic cell's capacity for genetic novelty set the stage for the invention of sexual reproduction (sv-invention-of-sex), which shuffles eukaryotic genomes and accelerates adaptation. It carried life through the planetary trauma of Snowball Earth (sv-snowball-earth) and bloomed, once the ice retreated, into the soft-bodied forms of the Ediacaran biota (sv-ediacaran-biota) and the riotous body plans of the Cambrian explosion (sv-cambrian-explosion). Every animal lineage thereafter — the first sharks, the first insects, the lobe-finned fish whose descendants made the move to land at Tiktaalik (sv-tiktaalik), and ultimately the first mammals (sv-first-mammals) — is built of eukaryotic cells respiring through captured bacteria.
The lineage runs unbroken to the reader. The first primates (sv-first-primates), the human-chimpanzee split (sv-human-chimp-split), and the civilizations that built Göbekli Tepe (sv-gobekli-tepe) and invented agriculture (sv-agriculture) are all eukaryotic enterprises, each cell still humming with mitochondria descended from that ancient guest. There is a quiet symmetry here that resonates across this whole timeline: the great leaps of history are often mergers — separate things combining into something neither could be alone. A host and a bacterium two billion years ago; ideas, machines, and minds today. Complex life began not with competition but with an act of permanent, world-making cooperation.
Global Context
Eukaryogenesis unfolded across the Proterozoic Eon, a planet dominated entirely by microbes. Its deeper backdrop was the Great Oxidation Event (~2.4 billion years ago), when cyanobacterial photosynthesis flooded the atmosphere with oxygen, poisoning anaerobes and reshaping ocean chemistry. The subsequent "Boring Billion" (roughly 1.8-0.8 Ga) was a stretch of geochemical and evolutionary stasis: stratified, often sulfidic oceans (the Canfield ocean model), stable continents, and low atmospheric oxygen. Within this seemingly quiet interval, the eukaryotic cell assembled. Acritarchs and the multicellular alga Bangiomorpha pubescens (~1.05 Ga) and the contested Gabonionta and Grypania appear in the fossil record. There were no animals, plants, or fungi; no Cambrian explosion would come for another half-billion years. The relevant "events" elsewhere were chemical and microbial: the establishment of the marine sulfur cycle, banded iron formation deposition tapering off, and the slow oxygenation that made aerobic respiration—and thus mitochondrial energetics—possible. The whole drama played out invisibly, at the scale of single cells in Proterozoic seas.
The Paradigm Shift
Eukaryogenesis was arguably the single most consequential transition in cellular evolution, and it reframed how biologists understand novelty itself. Before, the tree of life looked dichotomously branching, with complexity emerging gradually. Lynn Margulis's 1967 endosymbiotic theory—and its molecular vindication—established that the eukaryotic cell is a chimera, a fusion of distinct lineages: an archaeal host and a bacterial (alphaproteobacterial) endosymbiont, plus, in algae and plants, a cyanobacterial plastid. This made symbiosis, not merely mutation and selection, a primary engine of macroevolutionary innovation. The mitochondrion's energetic windfall—internalized bioenergetic membranes—plausibly underwrote the eukaryotic capacity for large genomes, vast cell volumes, the nucleus, endomembrane systems, and ultimately multicellularity, sex, plants, and animals. Nick Lane and William Martin (2010) argued bioenergetics, not information, was the rate-limiting bottleneck. Conceptually, eukaryogenesis dissolved the prokaryote/eukaryote dichotomy into a "two domains" picture in which eukaryotes branch from within Archaea, overturning Carl Woese's classic three-domain tree and recasting human ancestry as fundamentally archaeal-bacterial.
Counterfactual: What If It Had Gone Differently
Had the archaeal-bacterial endosymbiosis never occurred, Earth would plausibly have remained a microbial planet indefinitely. Nick Lane and William Martin (Nature, 2010) argue that prokaryotes are bioenergetically constrained: their energy-converting membrane is the cell surface, so genome and proteome expansion is penalized by surface-to-volume scaling. Only by internalizing respiratory membranes—mitochondria—could a cell escape this ceiling and afford the thousands-fold larger genomes that complex morphology requires. On this view, the endosymbiotic event was a singular, low-probability bottleneck: it apparently happened only once in four billion years, and without it, large complex cells, and therefore animals, plants, fungi, and intelligent life, would be effectively impossible. Critics (e.g., Booth and Doolittle) counter that the bioenergetic argument may overstate the constraint and that the singularity could reflect contingency rather than deep necessity. Either way, the counterfactual is stark: no eukaryogenesis, no Cambrian explosion, no terrestrial ecosystems, no observers. This logic also feeds astrobiology's "hard step" models of why complex life may be rare cosmically.
Scholarly Debate
A genuine, active dispute concerns the order of events—"mitochondria-early" versus "mitochondria-late"—and the nature of the host. Symbiosis-first models, championed by William Martin and Miklós Müller's hydrogen hypothesis (Nature, 1998) and elaborated by Nick Lane, hold that an archaeon acquired the bacterial endosymbiont before evolving eukaryotic complexity; the mitochondrion was the trigger, and phagocytosis came afterward. Autogenous, "complexity-first" models (associated with Tom Cavalier-Smith and others) hold that an already-elaborate, phagocytic proto-eukaryote engulfed the bacterium late. The 2015 discovery of Asgard archaea (Lokiarchaeota) by Thijs Ettema's group, encoding eukaryotic signature proteins, plus Hiroyuki Imachi's 2020 cultivation of Promethearchaeum syntrophicum (MK-D1) and its "Entangle-Engulf-Endogenize" (E3) model, strengthened the archaeal-host, syntrophy-based picture. Debate persists over whether eukaryotes branch within Asgard (two-domains, Eocyte hypothesis) or sister to it, and over phagocytosis's timing. The data increasingly favor an archaeal host and metabolic symbiosis, but the precise mechanism remains genuinely unsettled.
How It Connects
What Made It Possible
- Billions of years of prokaryotic life had already invented the core biochemistry of cells, including DNA-based heredity, ribosomal protein synthesis, and lipid membranes, supplying the molecular toolkit that eukaryotes would inherit and elaborate.
- The Asgard archaea (the closest known archaeal relatives of eukaryotes) evolved a repertoire of 'eukaryotic signature proteins' governing membrane remodeling and cytoskeletal dynamics, and recent phylogenomic work finds Asgard lineages made the dominant contribution to the conserved gene systems of the eukaryotic ancestor.
- Cyanobacteria evolved oxygenic photosynthesis and drove the Great Oxidation Event around 2.4 to 2.3 billion years ago, raising atmospheric oxygen and creating an environment in which aerobically respiring bacteria could thrive.
- Free-living alphaproteobacteria evolved efficient aerobic respiration (oxidative phosphorylation), making them metabolically valuable partners once they could be acquired by a host cell.
- An archaeal host lineage entered a close metabolic partnership with a bacterial cell, and rather than digesting it the host retained the alphaproteobacterial endosymbiont that would become the mitochondrion, a singular merger event reconstructed in the ancestry of all known eukaryotes.
- The internalized mitochondrion enabled oxidative phosphorylation across a large area of folded internal membrane, which according to the Lane and Martin bioenergetic hypothesis released cells from an energetic constraint on genome size and allowed a vast expansion in the number of genes that could be expressed.
Its Legacy
- The energy supplied by mitochondria underwrote large, gene-rich genomes and bigger cells, allowing the eukaryotic lineage to accumulate the regulatory and structural complexity that prokaryotes never achieved.
- Eukaryotes evolved a membrane-bound nucleus and an elaborate endomembrane system (endoplasmic reticulum, Golgi, vesicle trafficking), separating transcription from translation and enabling sophisticated protein sorting and modification.
- A dynamic actin-and-tubulin cytoskeleton arose, enabling phagocytosis, intracellular transport, and the directed cell movement that defines eukaryotic cells.
- Meiosis and sexual reproduction emerged, with the fusion of haploid gametes and genetic recombination shuffling genomes and greatly increasing the genetic diversity available to natural selection.
- A second endosymbiosis captured a cyanobacterium to form the plastid (chloroplast) over a billion years ago, founding the photosynthetic eukaryotes (glaucophytes, red and green algae, and ultimately land plants), with later secondary endosymbioses spreading photosynthesis across much of algal diversity.
- Eukaryotic cellular complexity provided the foundation for the independent, repeated evolution of complex multicellularity, giving rise to animals, plants, fungi, and the macroscopic biosphere, including humans.
Myth vs. Reality
Myth: Eukaryotes are just an 'advanced' or 'upgraded' version of bacteria, sitting one rung higher on a ladder of increasing complexity.
Reality: Eukaryotes are not descended from bacteria along a simple progression. The leading evidence indicates the eukaryotic host lineage arose from within the archaea, the third domain of life, and is a chimera: an archaeal host that merged with a bacterial (alphaproteobacterial) partner that became the mitochondrion. The discovery of Asgard archaea, whose genomes carry 'eukaryotic signature proteins' for cytoskeleton, membrane trafficking, and the ubiquitin system, supports a symbiogenetic origin rather than a linear bacteria-to-eukaryote climb.
Myth: A fully formed eukaryotic cell, already complete with a nucleus, simply ate a bacterium one day and that bacterium became the mitochondrion.
Reality: The host that acquired the mitochondrion was almost certainly not yet a modern eukaryote with a nucleus. Whether mitochondria came before or after features like the nucleus, endomembrane system, and phagocytosis is genuinely unresolved (the 'mitochondrion-early' vs 'mitochondrion-late' debate). Many models hold that the host was a relatively simple archaeal cell and that classic phagocytosis (engulfing prey) is energetically expensive and may have post-dated, not enabled, mitochondrial acquisition. The popular 'eukaryote ate a bacterium' image oversimplifies an open scientific question.
Myth: Endosymbiosis is a routine evolutionary trick that has produced complex cells many separate times.
Reality: All living eukaryotes trace back to a single last eukaryotic common ancestor (LECA) that already possessed a mitochondrion, meaning the foundational endosymbiosis appears to have succeeded essentially once in roughly four billion years of life. Prokaryote-engulfing-prokaryote endosymbiosis is otherwise almost unknown in nature, and researchers describe the FECA-to-LECA transition as passing through a severe bottleneck with no surviving intermediates. This singularity is precisely why eukaryogenesis is considered one of the rarest and most consequential events in the history of life, not a repeatable process.
Myth: Complex cells appeared promptly as soon as Earth's atmosphere filled with oxygen during the Great Oxidation Event.
Reality: There is a large temporal gap. The Great Oxidation Event raised atmospheric oxygen roughly 2.4 billion years ago, but many molecular-clock estimates place LECA hundreds of millions of years, by some estimates up to a billion years, later. This decoupling led some scientists to argue the origin of eukaryotes was not directly driven by oxygen at all. Even where newer estimates (around 2.0 to 1.8 billion years ago) bring the two closer, the long lag through the so-called 'Boring Billion' shows eukaryotic complexity did not follow oxygenation immediately or automatically.
Myth: We have a precise date for when the first complex cell appeared.
Reality: The timing is bracketed by uncertainty, not pinned down. Microfossil and biomarker evidence points to a broad window for the origin and early diversification of eukaryotes, often cited as roughly 1.6 to 2.2 billion years ago, while molecular-clock estimates vary considerably depending on calibration assumptions. The fossil record of early eukaryotes is sparse and contested, so any single headline date should be read as a midpoint of an actively debated range rather than an established fact.
In Their Words
"Life did not take over the globe by combat, but by networking." — Lynn Margulis and Dorion Sagan, Microcosmos: Four Billion Years of Microbial Evolution (1986)
Data Visualization
References & Sources
- Eukaryogenesis — Wikipedia
- Lynn Sagan (Margulis), 'On the Origin of Mitosing Cells,' Journal of Theoretical Biology 14 (1967): 225-274
- William Martin and Miklós Müller, 'The hydrogen hypothesis for the first eukaryote,' Nature 392 (1998): 37-41
- Nick Lane and William Martin, 'The energetics of genome complexity,' Nature 467 (2010): 929-934
- Hiroyuki Imachi et al., 'Isolation of an archaeon at the prokaryote-eukaryote interface,' Nature 577 (2020): 519-525
- Anja Spang, Thijs J. G. Ettema et al., 'Complex archaea that bridge the gap between prokaryotes and eukaryotes,' Nature 521 (2015): 173-179