The Rise of the Great Apes
Our tailless, big-brained ancestors diverge from the gibbons (the lesser apes).
Bodies Built for Branches: The Great Ape Experiment
The great apes did not arrive as a triumph. They arrived as one ambitious variation on a body plan that the planet had been refining for hundreds of millions of years, and for most of their history they were a fading lineage rather than an ascendant one. To see them clearly is to see deep time converge: every constraint that shaped them was inherited, and almost everything we are was first sketched in their bones.
The Inheritance
The great apes are unthinkable without the long chain that preceded them. Their warm, large-brained, fast-metabolizing bodies run on oxygen that only became abundant after the Great Oxygenation Event (sv-great-oxygenation) poisoned the early world and remade its chemistry. Their cells are the compartmentalized, mitochondria-powered eukaryotic kind first assembled by the first complex cells (sv-first-complex-cells), and they reproduce through the genetic shuffling pioneered by the invention of sexual reproduction (sv-invention-of-sex)—the engine that lets variation accumulate fast enough for an ape lineage to diversify at all. Their grasping skeleton is a refinement of the four-limbed vertebrate frame that Tiktaalik (sv-tiktaalik) first dragged onto land, and their nursing, parental, warm-blooded biology descends directly from the first true mammals (sv-first-mammals). Most immediately, they are the inheritors of the first primates (sv-first-primates), arboreal mammals whose forward-facing eyes and dexterous hands were honed for life in the canopy.
The Miocene Moment
Apes proper emerged in Africa near the Oligocene-Miocene boundary, with stem forms like Proconsul by roughly eighteen million years ago. The crucial development was an evolutionary radiation: by the Middle Miocene, apes expanded out of Africa into Eurasia, producing twenty or more genera spanning a wide range of sizes. Dryopithecus spread through Europe and Sivapithecus through Asia, the latter widely regarded as a forebear of the orangutan. These animals already grew slowly, had life histories like living apes, and—as Dryopithecus braincases show—carried brains comparable to a chimpanzee's. The molecular clock tracks the splitting that followed: orangutans diverged perhaps twelve to sixteen million years ago, gorillas later still, setting the stage for the human-chimpanzee split (sv-human-chimp-split) within the last several million years.
This was a forest empire, and it did not last. Late Miocene mountain-building in the Alps, Himalayas, and East Africa, shifting ocean currents, and the first polar ice precipitated the Asian monsoon, dried East Africa, and cooled Europe. Most Eurasian great apes went extinct. The lineage that survived did so by adapting to the patchy, drying woodlands left behind—pressure that eventually drove some apes out of the trees entirely.
The Ripple Forward
Everything downstream of the human story is, biologically, a continuation of the great ape experiment. The slow-growing, large-brained body that survived the Miocene collapse became the platform for tool-using, fire-keeping, symbol-making hominins. From there the thread runs unbroken through the Last Ice Age (sv-last-ice-age) that culled the megafauna our ancestors hunted, to the first monumental religion at Göbekli Tepe (sv-gobekli-tepe), to the Agricultural Revolution (sv-agriculture) that settled us in place. The hands that AlphaGo's engineers used to type, the brains that conceived the Transformer (sv-transformer-paper)—all are the slowly-grown great ape body, still running on Miocene wiring. It took Charles Darwin (sv-charles-darwin) to recognize that these dwindling forest creatures and ourselves were one continuous lineage. The great apes were never destined to inherit the Earth. One marginal branch of them, against the odds, simply did not die.
Global Context
The great-ape radiation unfolded across the Miocene Epoch (23–5.3 Ma), a world warmer and wetter than today. Its pivot was the Middle Miocene Climatic Optimum (c. 17–14.75 Ma), when global mean temperatures peaked roughly 3°C above present and subtropical forests stretched deep into Eurasia. This warmth opened a forested corridor—amplified by the Early-Miocene closure of the Tethys and the c. 19 Ma Gomphotherium land bridge linking Afro-Arabia to Eurasia—across which apes dispersed out of Africa. The same epoch saw proboscideans radiate, bovids and equids diversify on emerging grasslands, and, after the optimum, pronounced cooling with Antarctic ice expansion and the spread of savannas. Old World monkeys (cercopithecoids), initially rarer than apes, were beginning the rise that would later eclipse hominoid diversity. Continents had nearly reached modern positions; the Himalayas and Alps were rising. In this dynamic, forest-rich Miocene world—long before any hominin—apes were the dominant primates, a true "planet of the apes" spanning Spain to China.
The Paradigm Shift
The Miocene radiation assembled the great-ape body plan that every later hominin inherited—including ourselves. Early African forms such as Proconsul (c. 18 Ma) were tailless but largely above-branch arboreal quadrupeds lacking modern suspensory specializations. The decisive innovation was the orthograde (upright-trunked) torso, first comprehensively documented in Pierolapithecus catalaunicus (c. 12 Ma, Spain; Moyà-Solà et al. 2004), whose stiffened lower back, broad shallow thorax, and mobile wrists enabled vertical climbing and clambering. This orthograde scaffold—not knuckle-walking or bipedalism—is the structural prerequisite from which hominin upright posture was later exapted. The radiation also fixed the deep phylogenetic architecture of living hominoids: molecular clocks place the orangutan lineage's divergence in the Middle-to-Late Miocene and the gorilla and chimpanzee splits in the Late Miocene. By generating the crown-hominoid morphotype and seeding the lineages leading to orangutans, gorillas, chimpanzees, and humans, the Miocene ape diversification set the entire trajectory of subsequent primate—and ultimately human—evolution.
Counterfactual: What If It Had Gone Differently
Had the Miocene forests not sustained a sprawling ape radiation, the hominin lineage would have had no platform from which to emerge. The counterfactual is constrained by ecology: cercopithecoid monkeys, with their efficient quadrupedalism and ability to digest unripe and toxic foliage (notably colobines), progressively outcompeted apes as Late-Miocene cooling fragmented forests. Andrews and others argue that dietary specialization and habitat loss drove the extinction of European great apes after c. 9 Ma; absent the earlier orthograde innovation, apes might have vanished entirely, leaving monkeys as the Old World's dominant anthropoids and no orthograde scaffold for bipedalism. Conversely, had the Eurasian radiation never occurred, Begun's "back to Africa" model—in which the African ape-human clade descends from a Eurasian dryopithecine that re-entered Africa 6–9 Ma—would be impossible, and the geography of human origins would differ. The radiation was thus a genuine bottleneck-and-opportunity: remove it, and the most plausible outcome is a primate world of monkeys without hominins, a conclusion supported by the documented Late-Miocene ape contraction to equatorial refugia.
Scholarly Debate
The sharpest live debate concerns the geographic cradle of the great-ape-and-human clade (Hominidae/Homininae). David Begun and colleagues advance a Eurasian-origin, "back to Africa" model: Eurasian dryopithecines (e.g., Dryopithecus, Rudapithecus, Ouranopithecus) include the ancestry of African apes and humans, who re-dispersed into Africa in the latest Miocene. Against this, a Darwinian African-origin school—long defended by Peter Andrews and reinforced by Egyptian finds such as the newly described Masripithecus (2026), which is placed closer to crown hominoids than coeval East African apes—locates the relevant common ancestor in northern Africa or Arabia. A second controversy surrounds Pierolapithecus: Moyà-Solà et al. read it as an early crown great ape documenting orthograde origins, while critics (Begun, Ward) question whether it is a crown hominid at all rather than a stem form, and dispute its locomotor reconstruction. Underlying both is a methodological rift—how far molecular divergence dates should override, or be calibrated by, a fragmentary and homoplasy-riddled fossil record (Alba; Pugh).
How It Connects
What Made It Possible
- The Catarrhini lineage emerged in Afro-Arabia during the late Eocene and Oligocene, exemplified by stem catarrhines such as Aegyptopithecus zeuxis (roughly 38-29.5 million years ago) from Egypt's Fayum Jebel Qatrani Formation, establishing the ancestral stock from which both Old World monkeys and apes would descend.
- Combined morphological and molecular evidence places the divergence of hominoids (apes) from cercopithecoids (Old World monkeys) at roughly 29.6 million years ago in Afro-Arabia, splitting the ape lineage onto its own evolutionary trajectory.
- Early Miocene Africa functioned as an island continent whose warm, wet, multistoried closed-canopy tropical forests, such as those reconstructed on Kenya's Rusinga Island, provided the arboreal habitat in which a diverse cohort of early apes radiated.
- Loss of the external tail, inferred to have occurred around the time the hominoid lineage diverged from Old World monkeys (about 25 million years ago) and leaving only a few coccygeal vertebrae, removed a balancing organ used in quadrupedal branch-walking and helped reorient the ape lineage toward upright, climbing-based postures.
- Early proconsulid apes such as Proconsul (and related forms now placed in Ekembo, Afropithecus, and Turkanapithecus), known from exceptionally complete fossils in Kenya and Uganda dating from roughly 23 to 14 million years ago, established the tailless, larger-bodied catarrhine bauplan on which later great-ape specializations were built.
- The Mid-Miocene Climatic Optimum, a global warming peak around 17-15 million years ago, expanded forested habitat and is associated with the diversification of Miocene apes into as many as roughly 50 fossil genera across Africa, Europe, and Asia.
Its Legacy
- The orthograde (upright-torsoed) body plan with a broad, flattened thorax evolved in a stepwise, mosaic fashion across Miocene apes, with forms like the Middle Miocene Pierolapithecus catalaunicus and the Late Miocene Hispanopithecus laietanus documenting enhanced vertical climbing and the oldest unambiguous suspensory adaptations that define living great apes.
- After their initial African radiation, apes dispersed into Eurasia by roughly 14 million years ago amid shifting tectonics and climate, diversifying into great-ape genera such as Dryopithecus in Europe and Sivapithecus in Asia.
- Sivapithecus from the Indo-Pakistani region preserves orangutan-like cranial features, supporting hypotheses that the Asian great-ape (Pongo) lineage traces back to Late Miocene Eurasian apes.
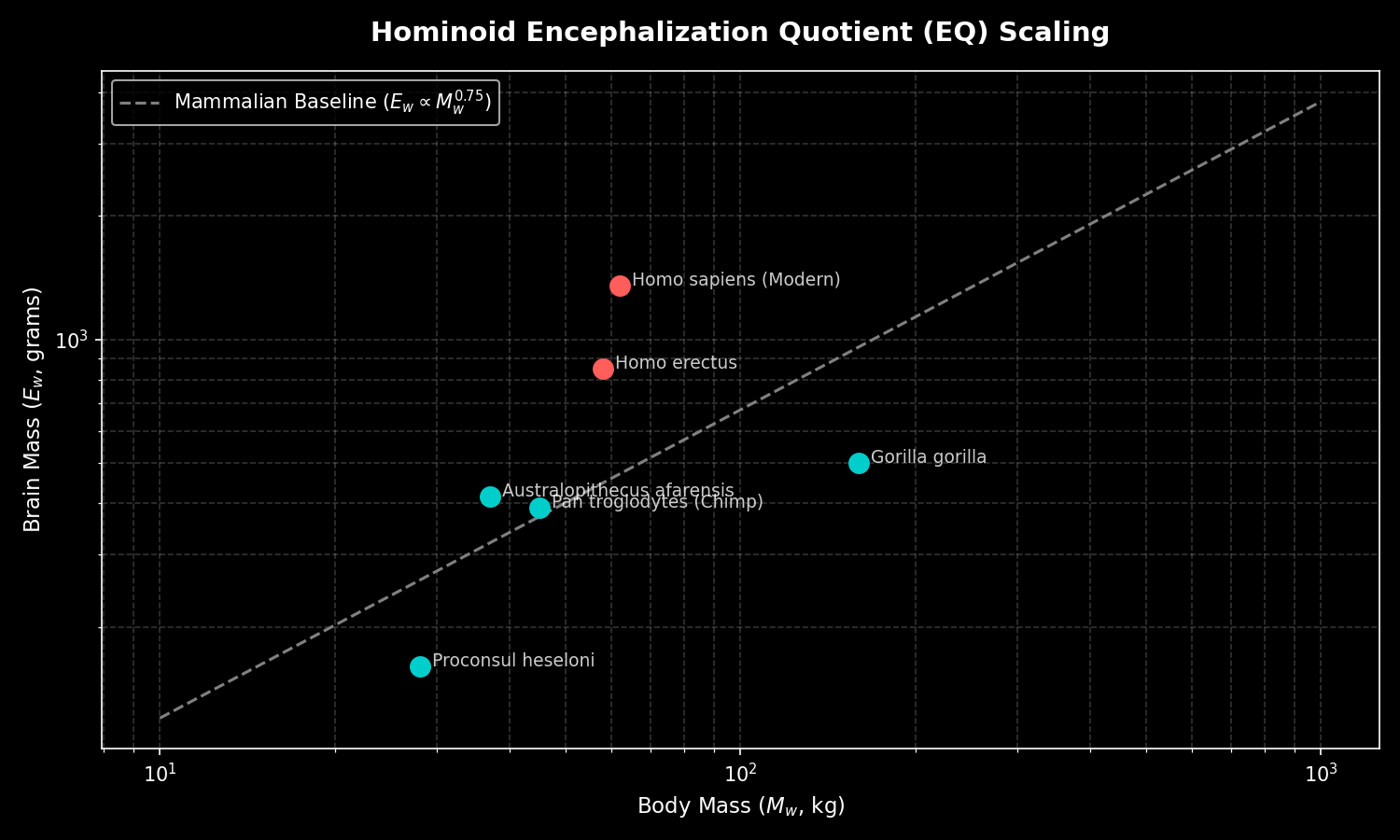
- A significant Miocene increase in encephalization marked the emergence of the great-ape-and-human clade (Hominidae), and because mid-to-late Miocene apes already had brains of comparable size to living great apes, capacities such as symbolic cognition, innovation, and cultural transmission may have been potentiated as early as 12-14 million years ago.
- The Middle Miocene climate transition's stepwise cooling and drying, combined with the dietary specializations of Western Eurasian apes, ultimately drove the extinction of the European great apes as forests contracted, leaving the African lineages to carry the clade forward.
- Within the African hominine lineage, gorillas diverged first and then the human (hominin) and chimpanzee (panin) lines split late in the Miocene roughly 6-8 million years ago, a window represented by candidate early hominins such as the approximately 7-million-year-old Sahelanthropus tchadensis (Toumaï) from Chad, though its hominin status remains debated.
Myth vs. Reality
Myth: Humans evolved from chimpanzees (or from any living ape), so the 'rise of the great apes' was a ladder climbing toward us.
Reality: Humans did not descend from chimpanzees; the two lineages split from a common, now-extinct ancestor roughly 6-8 million years ago, and each evolved independently afterward. Molecular comparisons show humans and chimps share around 98-99% of their DNA, with the differences accumulating after the split. Great-ape evolution is a branching bush, not a directional ladder pointed at Homo sapiens, which is why chimpanzees still exist alongside us rather than having 'turned into' humans.
Myth: Great apes are a small, primitive group that has always been marginal compared to monkeys and humans.
Reality: During the Miocene (roughly 23-5 million years ago), apes were spectacularly diverse, with researchers estimating as many as 100 ape species ranging across the Old World, from France to China and from Kenya to Namibia. The living great apes (gibbons, orangutans, gorillas, chimps, bonobos, and humans count as hominoids/apes) are a relict remnant of that vanished radiation. By about 7 million years ago apes had gone extinct in Europe while surviving lineages in Asia and Africa gave rise to the modern forms.
Myth: Ramapithecus was confirmed as the earliest human ancestor, an established fact of the great-ape story.
Reality: From the 1960s into the 1970s, fossils named Ramapithecus (studied by Elwyn Simons and David Pilbeam) were widely promoted as the first hominin on the line to humans, based on jaw and tooth shape. A more complete jaw Pilbeam described in 1976 showed a V-shaped (not human-like parabolic) dental arcade, and molecular-clock evidence pointing to a much later human-ape split contradicted the old timeline. By the early 1980s Ramapithecus was reinterpreted as part of Sivapithecus, an ancestor or close relative of the orangutan, not of humans.
Myth: Human bipedalism evolved straightforwardly from a knuckle-walking great-ape ancestor like a gorilla or chimp.
Reality: This 'knuckle-walking origin' idea is contested. Kivell and Schmitt (2009) argued that knuckle-walking features evolved independently in gorillas and in chimps/bonobos, implying the last common ancestor of humans and African apes was likely not a committed knuckle-walker. Fossils such as Danuvius guggenmosi (about 11.6 million years old, described by Madelaine Bohme and colleagues in 2019) suggest some Miocene apes combined upright walking with arboreal climbing, supporting the view that bipedal tendencies may have arisen in trees rather than from a ground-based knuckle-walking stage.
Myth: The rise of the great apes hinges on finding the single 'missing link' between apes and humans.
Reality: The phrase 'missing link' is a popular-media term, not a scientific one, and it misleads by implying a linear chain with one pivotal gap. Anthropologists prefer 'last common ancestor' because evolution branches like a bush, and most fossil apes are side-branches and evolutionary dead ends rather than direct ancestors. Rather than one elusive link, scientists work with many transitional fossils and independent lines of evidence; the National Center for Science Education and UC Berkeley's evolution resources both stress that no single missing link is expected or needed.
In Their Words
"It is therefore probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man's nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere." — Charles Darwin, The Descent of Man, and Selection in Relation to Sex (1871), vol. 1, ch. 6
Data Visualization
References & Sources
- Hominidae — Wikipedia
- Moyà-Solà, S., Köhler, M., Alba, D. M., Casanovas-Vilar, I., & Galindo, J. (2004). 'Pierolapithecus catalaunicus, a New Middle Miocene Great Ape from Spain.' Science 306: 1339–1344.
- Begun, D. R. (2010). 'Miocene Hominids and the Origins of the African Apes and Humans.' Annual Review of Anthropology 39: 67–84.
- Andrews, P. (2020). 'Last Common Ancestor of Apes and Humans: Morphology and Environment.' Folia Primatologica.
- Charles Darwin (1871). The Descent of Man, and Selection in Relation to Sex. London: John Murray.
- Alba, D. M. (2012). 'Fossil Apes from the Vallès-Penedès Basin.' Evolutionary Anthropology 21: 254–269.